
Are there optical communication channels in the brain?
Parisa Zarkeshian, Sourabh Kumar, Jack Tuszynski, Paul Barclay, Christoph Simon (Submitted on 23 Aug 2017)
Despite great progress in neuroscience, there are still fundamental unanswered questions about the brain, including the origin of subjective experience and consciousness. Some answers might rely on new physical mechanisms. Given that biophotons have been discovered in the brain, it is interesting to explore if neurons use photonic communication in addition to the well-studied electro-chemical signals. Such photonic communication in the brain would require waveguides. Here we review recent work [S. Kumar, K. Boone, J. Tuszynski, P. Barclay, and C. Simon, Scientific Reports 6, 36508 (2016)] suggesting that myelinated axons could serve as photonic waveguides. The light transmission in the myelinated axon was modeled, taking into account its realistic imperfections, and experiments were proposed both in-vivo and in-vitro to test this hypothesis. Potential implications for quantum biology are discussed.
| Comments: | 13 pages, 5 figures, review of arXiv:1607.02969 for Frontiers in Bioscience, updated figures, new references on existence of opsins in the brain and experimental effects of light on neurons |
| Subjects: | Biological Physics (physics.bio-ph); Optics (physics.optics); Neurons and Cognition (q-bio.NC); Quantum Physics (quant-ph) |
| Cite as: | arXiv:1708.08887 [physics.bio-ph] |
| (or arXiv:1708.08887v1 [physics.bio-ph] for this version) |
Submission history
From: Parisa Zarkeshian
[v1] Wed, 23 Aug 2017 22:54:52 UTC (1,981 KB)
1. ABSTRACT
Despite great progress in neuroscience, there are still fundamental unanswered questions about the brain, including the origin of subjective experience and consciousness. Some answers might rely on new physical mechanisms. Given that biophotons have been discovered in the brain, it is interesting to explore if neurons use photonic communication in addition to the well-studied electro-chemical signals. Such photonic communication in the brain would require waveguides. Here we review recent work [S. Kumar, K. Boone, J. Tuszynski, P. Barclay, and C. Simon, Scientific Reports 6, 36508 (2016)] suggesting that myelinated axons could serve as photonic waveguides. The light transmission in the myelinated axon was modeled, taking into account its realistic imperfections, and experiments were proposed both in vivo and in vitro to test this hypothesis. Potential implications for quantum biology are discussed.
2. INTRODUCTION
Over the past decades a substantial number of facts has been discovered in the field of brain research. However, the fundamental question of how neurons, or more specifically all particles involved in the biological processes in the brain, contribute to mental abilities such as consciousness is still unanswered. The true explanation to this question might rely on physical processes other than those that have been discovered so far. One interesting candidate to focus on is biophotons, which might serve as supplementary information carriers in the brain in addition to the well established electro-chemical signals.
Biophotons – which are photons ranging from near-IR to near-UV frequency and emitted without any enhancement or excitation– have been observed in many organisms such as bacteria (1), fungi (2), germinating seeds (3), plants (4), animal tissue cultures (5), and different parts of the human body (6–9), including the brain (10–15). These biophotons are produced by the decay of electronically excited species which are created chemically during oxidative metabolic processes (16, 17) and can contribute to communication between cells (18). Moreover, several experimental studies show the effects of light on neurons’ and, generally, the brain’s function (19–21). The existence of biophotons and their possible effects on the the brain along with the fact that photons are convenient carriers of information raises the question whether there could be optical communication in the brain.
For the sources and detectors of the optical communication process in the brain, mitochondrial respiration (22, 23) or lipid oxidation (24), and centrosomes (25) or chromophores in the mitochondria (26) have been proposed, respectively. It has also been observed that opsins, photoreceptor protein molecules, exist in the brains of birds (27, 28), mammals (29–32), and more general vertebrates (33) and even in other parts of their bodies (34, 35) as well.
Another essential element for this optical communication, which is not well established yet, is the existence of physical links to connect all of these spatially separated agents in a selective way. In the dense and (seemingly) disordered environment of the brain, waveguide channels for traveling photons would be the only viable way to achieve the targeted optical communication processes. Mitochondria and microtubules in neurons have been introduced as the candidates for such waveguides (36–39). How- ever, they are not suitable in reality due to their small and inhomogeneous structure for light guidance over proper distances in the brain.
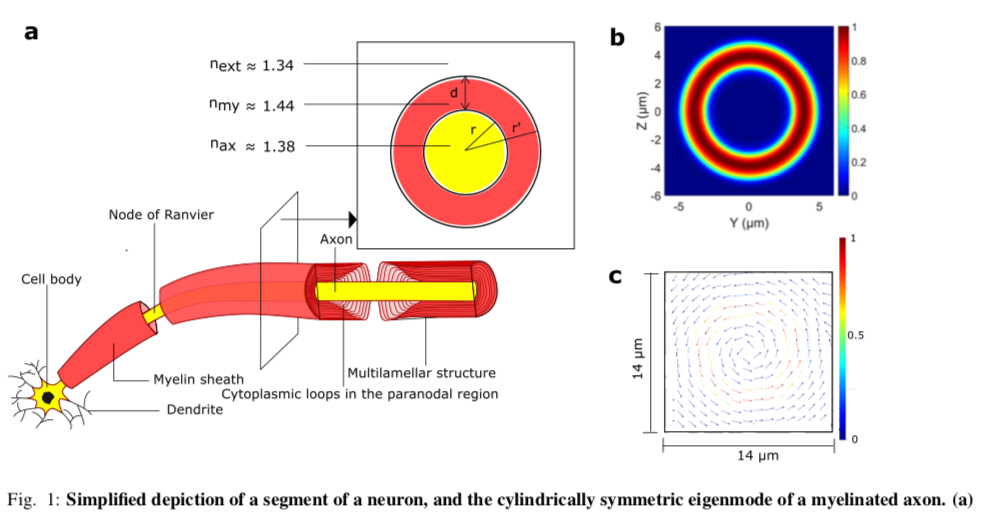
Ref. (40) proposed myelinated axons as potential biophoton waveguides in the brain. The proposal is supported by a theoretical model and numerical results taking into account real imperfections. Myelin sheath (formed in the central nervous system by a kind of glia cell called oligodendrocyte) is a lamellar structure surrounding the axon and has a higher refractive index (41) than both the inside of the axon and the interstitial fluid outside (see Fig. 1a) which let the myelin sheath to guide the light inside itself for optical communications. This compact sheath also increases the propagation speed of an action potential (via saltatory conduction) based on its insulating property (42). There has been a few indirect experimental evidence for light conduction by axons (12, 43, 44). Another related and interesting experiment has shown that a certain type of glia cells, known as Mu ̈ller cells, guide light in mammalian eyes (45, 46). Ref. (40) also proposed experiments to test the existence of the optical waveguides in the brain.
One interesting property of optical communication channels is that they can also transmit quantum information. Quan- tum effects in biological systems are being studied in different areas such as photosynthesis (47, 48), avian magnetoreception (49, 50), and olfaction (51, 52). There is an increasing number of conjectures about the role of quintessential quantum features such as superposition and entanglement (53) in the brain (15, 38, 54–56). The greatest challenge when considering quantum effects in the brain or any biological system in general is environmentally induced decoherence (57), which leads to the suppression of these quantum phenomena. However, some biological processes can be fast and may show quantum features before they are destroyed by the environment. Moreover, nuclear spins can have coherence times of tens of milliseconds in the brain (58, 59). A recent proposal on “quantum cognition” suggests even longer coherence times of nuclear spins (56), but relies on quantum information transmission via molecule transport, which is very slow. In contrast, photons are the fastest and most robust carriers for quantum information over long distances, which is why currently man-made quantum networks rely on optical communication channels (typically optical fibers) between spins (60, 61).
3. Results
To show that myelinated axons could serve as the waveguides for traveling biophotons in the brain, Ref. (40) solved the three dimensional electromagnetic field equations numerically in different conditions, using Lumerical’s software packages FDTD (Finite Difference Time Domain) Solutions and MODE Solutions. These software packages solve Maxwell’s equations numerically, allowing the optical properties of dielectric structures defined over a mesh with subwavelength resolution to be simulated.
The refractive indices of the fluid outside of the axon, the axon, and the myelin sheath were taken close to 1.34, 1.38 and 1.44 respectively (see Fig. 1a), which are consistent with their typical values (41, 62, 63). These indexes let the myelin sheath guide the light inside itself. The ratio of the radius of the axon, r to the outer radius of the myelin sheath r′ (g-ratio) is taken equal to 0.6 for the most of the simulations, close to the experimental values (64). In reality, the radius of the myelinated axons in the brain changes from 0.2 microns to close to 10 microns (65). For the purpose of guiding light inside the myelin sheath, Ref. (40) considered the wavelength of the observed biophotons in the brain which is from 200 nm to 1300 nm. Since several proteins in the environment of the axons strongly absorb at wavelengths close to 300nm, a wavelength range of the transmitted light from the shortest permissible wavelength, λmin = 400nm, to the longest one, λmax, was chosen to avoid the absorption and confine the light well in the myelin sheath. λmax is chosen to the upper bound of the observed biophoton wavelength (1300 nm) or the thickness of the myelin sheath (denoted by d), whichever is smaller. Besides λmin and λmax, an intermediate wavelength was considered, denoted by λint , corresponding to the central permissible frequency (mid-frequency of the permissible frequency range) in the simulations.
In the following section we discuss the guided modes in the myelinated axons and their transmissions in nodal and para- nodal regions and even in the presence of the imperfections such as bends, varying cross-sections, and non-circular cross-sections.
Optical communication channels in the brain